Η επεξεργασία σε ευκαρυώτες επηρεάζει όλους τους τύπους πρωτογενών μεταγραφών ευκαρυωτικών γονιδίων.
Επεξεργασία σε ευκαρυώτες
Κάλυψηαντιπροσωπεύει το σχηματισμό στο 5" άκρο του mRNA μιας ειδικής δομής - ένα καπάκι (καπάκι). Η κάλυψη συμβαίνει ακόμη και πριν από την πλήρη ολοκλήρωση της μεταγραφής και προστατεύει το άκρο 5" του RNA από τη δράση των νουκλεασών. Η κάλυψη RNA πραγματοποιείται με τη συμμετοχή GTP (τριφωσφορική γουανοσίνη), από την οποία η GMP μεταφέρεται στο 5"-διφωσφορικό του πρώτου νουκλεοτιδίου του mRNA.
Πολυαδενυλίωσηπραγματοποιείται από το ένζυμο πολυ(Α) πολυμεράση και οδηγεί στο σχηματισμό στο άκρο 3" ενός θραύσματος ολίγο(Α) που περιέχει 100 - 200 υπολείμματα αδενυλικού οξέος στη σειρά και ονομάζεται επίσης "ουρά πολυ(Α)". πολυ(Α) -υποακολουθία προστίθεται στο RNA μετά την προσάρτηση του καπακιού. Αρχικά, το άκρο 3" του RNA διασπάται από ένζυμα σε ένα σημείο 10-35 ριβονουκλεοτιδίων μακριά από τη συντηρητική αλληλουχία AAUAAA και στη συνέχεια λαμβάνει χώρα πολυαδενυλίωση αυτού του άκρου του μορίου RNA. Η ουρά πολυ(Α) βρίσκεται σχεδόν σε όλα Ευκαρυωτικοί οργανισμοί mRNA, με εξαίρεση τα μεταγραφήματα των γονιδίων ιστόνης Η αλληλουχία AAUAAA δεν βρίσκεται σε όλα τα ευκαρυωτικά RNA, προφανώς λόγω μεταλλάξεων που εμποδίζουν την πολυαδενυλίωση, τα μεταγραφήματα RNA αποικοδομούνται γρήγορα από τα ένζυμα.
Οτι. Το κάλυμμα 5" και η ουρά 3" είναι εξαιρετικά σημαντικά για την περαιτέρω επεξεργασία και μεταφορά του mRNA στο κυτταρόπλασμα. Η ουρά πολυ(Α) καθορίζει τη σταθερότητα του mRNA και τη διάρκεια ζωής του στο κύτταρο. Επιπλέον, προάγει την απελευθέρωση mRNA από τον πυρήνα στο κυτταρόπλασμα και είναι επίσης απαραίτητο για τη ρύθμιση της μετάφρασης.
Μηχανισμοί ματίσματος: αυτοκατάλυση RNA (Klag, 400)
Διαφορετικοί τύποι πυρηνικού RNA, καθώς και τα RNA mtx και chlp, έχουν τους δικούς τους μηχανισμούς ματίσματος.
Ανάλογα με την ιδιαιτερότητα του μηχανισμού ματίσματος, τα εσώνια μπορούν να χωριστούν σε διάφορες ομάδες. Στην πρώτη ομάδαΑυτά περιλαμβάνουν εσώνια που αποτελούν μέρος του πρωτεύοντος μεταγράφου rRNA, η απομάκρυνση των οποίων δεν απαιτεί πρόσθετα συστατικά. Αυτά τα ίδια τα εσώνια έχουν την ενζυματική δραστηριότητα που είναι απαραίτητη για την αποκοπή τους. Αυτό το γεγονός ανακαλύφθηκε για πρώτη φορά το 1982 (Tomas Cech et al.) στο μαστιγωτό πρωτόζωο Tetrachymena. Λόγω των αυτοκαταλυτικών ιδιοτήτων τους, μερικές φορές ονομάζονται αυτοσυνδυαζόμενα RNA ριβοένζυμα .
Διαδικασία αυτοκοπής (αυτόματη αποκοπή) (Εικ. 145_Konichev)
(Εικ. 12-12, Klag) αντιπροσωπεύει δύο πυρηνόφιλες αντιδράσεις ή αντιδράσεις μετεστεροποίηση, στην οποία η γουανοσίνη αλληλεπιδρά με το πρωτεύον μεταγράφημα και δρα ως συμπαράγοντας. Σε αυτή την περίπτωση, η 3"-υδροξυλομάδα της γουανοσίνης μεταφέρεται στο νουκλεοτίδιο δίπλα στο άκρο 5" του ιντρονίου. Στη δεύτερη αντίδραση, αυτή η ομάδα υδροξυλίου αλληλεπιδρά με μια φωσφορική ομάδα στο άκρο 3" του δεξιού ιντρονίου, με αποτέλεσμα το εσώνιο να αποκοπεί και τα άκρα δύο γειτονικών εξονίων να ενωθούν για να σχηματίσουν ώριμο mRNA.
Το εσώνιο 26S rRNA του Tetrahymena, IVS, αποτελείται από 413 νουκλεοτίδια. Ως αποτέλεσμα της αντίδρασης μετεστεροποίηση χωρίς πρόσθετη δαπάνη ενέργειας, δύο εξόνια συνδέονται για να σχηματίσουν ώριμο 26S rRNA. Το αποκομμένο εσώνιο στη συνέχεια κυκλοποιείται. Από τη σύνθεσή του, ένα θραύσμα που περιέχει 19 νουκλεοτίδια απελευθερώνεται με αυτοδιάσπαση δύο σταδίων, με αποτέλεσμα το σχηματισμό νουκλεοτιδίων μήκους RNA 376 (L -19 IVS), το οποίο είναι ένα πραγματικό ένζυμο RNA (ριβοένζυμο), το οποίο έχει καταλυτικές ιδιότητες. Αυτό το ριβοένζυμο έχει σταθερή δομή, έχει δραστηριότητα ενδονουκλεάσης, διασπά μακρό μονόκλωνο RNA και εμφανίζει ειδικότητα, αναγνωρίζοντας τετρανουκλεοτίδια CUCU στη σύνθεση του προσβεβλημένου υποστρώματος. Στη δομή εσώνια τύπου Ιέχουν ταυτοποιηθεί χαρακτηριστικές εσωτερικές αλληλουχίες ολιγοπουρίνης (στο Tetrahymena αυτή είναι η αλληλουχία GGAGGG), που ονομάζεται ακολουθίες προσαρμογέων , τα οποία συμμετέχουν στο σχηματισμό του ενεργού κέντρου των ενζύμων RNA και παίζουν κρίσιμο ρόλο στην καταλυτική διάσπαση του RNA.
Αυτή η αυτοεκτομή των ιντρονίων είναι χαρακτηριστική του προ-rRNA άλλων πρωτόζωων. Αυτός ο μηχανισμός προφανώς λειτουργεί επίσης κατά την αφαίρεση των ιντρονίων από τα πρωτεύοντα μεταγραφήματα mRNA και tRNA σε μιτοχόνδρια και χλωροπλάστες, που σχετίζονται με ομάδα II.
Για να κόψετε εσώνια δεύτερη ομάδαΑπαιτούνται επίσης δύο αυτοκαταλυτικές αντιδράσεις, αλλά δεν απαιτείται γουανοσίνη.
Περαιτέρω μελέτες αποκάλυψαν ότι όχι μόνο μεγάλα RNA (~400 νουκλεοτίδια στο Tetrahymena και RNase P), αλλά και βραχέα 13-20-μερή ολιγονουκλεοτίδια που μπορούν να συντεθούν in vitro έχουν καταλυτική δράση. Τέτοια ριβοένζυμα άρχισαν να ονομάζονται μίνι χειμώνες . Ένα από τα μοντέλα της λειτουργίας τέτοιων ριβοενζύμων που έχει μελετηθεί λεπτομερώς ονομάζεται «σφυρί κεφάλι «(Εικ. 146). Η τριτογενής δομή της «κεφαλής του σφυριού» σταθεροποιείται από δισθενή ιόντα μετάλλων, τα οποία εξουδετερώνουν τα αρνητικά φορτισμένα άτομα οξυγόνου των φωσφοδιεστερικών δεσμών και ταυτόχρονα συνδέουν φωσφορικές ομάδες με ομοιοπολικούς δεσμούς, οι οποίοι είναι απαραίτητοι για το σχηματισμό μιας σταθερής μεταβατικής κατάστασης (ένζυμο-υπόστρωμα συγκρότημα). Όπως στην περίπτωση της κατάλυσης που πραγματοποιείται από ένζυμα πρωτεΐνης, ριβοένζυμα και το υπόστρωμα που προσβάλλεται
(φυσικά ή συνθετικά παραγόμενα μόρια RNA) σχηματίζουν ένα σύμπλοκο ενζύμου-υποστρώματος και μετά ένα σύμπλοκο ενζύμου-προϊόντος (βλ. Εικ. 146).
Μηχανισμοί συναρμογής: συρματόσωμα. (Επεξεργασία MRNA σε ευκαρυώτες)
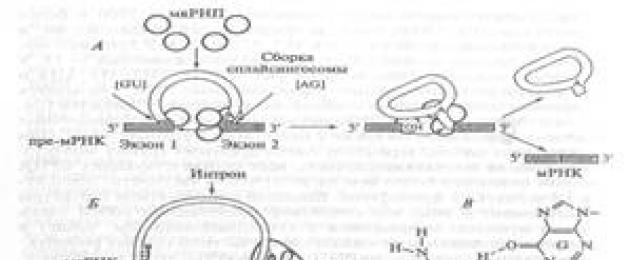
Στα πυρηνικά προ-mRNA, τα ιντρόνια μπορεί να έχουν μήκος έως και 20.000 νουκλεοτίδια. Επομένως, η αφαίρεσή τους απαιτεί πιο πολύπλοκο μηχανισμό από την αυτοεκτομή (αυτοεκτομή). (Εικ. 12-13). Οι αλληλουχίες νουκλεοτιδίων στα άκρα των εσωνίων σε αυτά τα μόρια είναι παρόμοιες: στα άκρα των 5" υπάρχει συχνά ένα δινουκλεοτίδιο (GU) GU, και στο άκρο 3" υπάρχει ένα δινουκλεοτίδιο (AG)ΑΓ. Μόρια ειδικών πρωτεϊνών συνδέονται με αυτές τις αλληλουχίες και σχηματίζουν ένα σύμπλεγμα που ονομάζεται splicome. Το κύριο συστατικό του ματοσώματος είναι μικρές πυρηνικές ριβονουκλεοπρωτεΐνες ή snRNPs, τα οποία βρίσκονται μόνο στον πυρήνα και είναι εμπλουτισμένα σε υπολείμματα ουριδίνης. Επομένως, τα μικρά πυρηνικά RNA συχνά ονομάζονται U1, U2...U6.
[Konichev, σελ.292.Σε προ-mRNA μάτισμα
Στους ανώτερους ευκαρυώτες εμπλέκεται ένας αριθμός πρωτεϊνών, καθώς και ένας ειδικός τύπος RNA - μικρό πυρηνικό RNA (snRNA). Τα μικρά πυρηνικά RNA έχουν αλληλουχίες που κυμαίνονται από 65 έως 1000 ή περισσότερα νουκλεοτίδια (10S-90S), πλούσια σε ουριδυλονουκλεοτίδια, και ως εκ τούτου ονομάζονται επίσης uRNA (Ul, U2, κ.λπ.). Στον ζυμομύκητα, έχουν εντοπιστεί 25 διαφορετικά snRNA, στα σπονδυλωτά - 15. Στους βατράχους με νύχια Xenopus laevis, ένας αριθμός snRNA (U3, U8, U14 και U22) εμπλέκονται στην επεξεργασία των ριβοσωμικών RNA, που συνδέονται με τις συνοριακές περιοχές του αλληλουχίες διαχωριστικών (βλ. Εικ. 143). Μικρά πυρηνικά RNA έχουν εντοπιστεί όχι μόνο σε σπονδυλωτά και ζυμομύκητες, αλλά και σε έντομα και αρχοβακτήρια. Είναι πιθανώς μια πολύ αρχαία ομάδα μορίων. Αλληλουχία νουκλεοτιδίων όλων των αντίστοιχων uRNA
Οι ευκαρυώτες συμπίπτουν κατά περισσότερο από 90%, το οποίο, ειδικότερα, ισχύει για τον άνθρωπο και τη Drosophila U1. Ο υψηλός συντηρητισμός της δομής του uRNA υποδηλώνει ότι το μάτισμα είναι μια πολύ αρχαία διαδικασία που ξεκίνησε με το αυτόματο μάτισμα (βλ. παραπάνω) και μετατράπηκε σε μάτισμα με τη συμμετοχή ειδικών σωματιδίων ριβονουκλεοπρωτεΐνης - snRNPs. Τα γονίδια SnRNA μεταγράφονται από την RNA πολυμεράση II και έχουν διαφορετικούς εντοπισμούς στο γονιδίωμα: μερικά από αυτά είναι διακριτά ανεξάρτητα γονίδια,
χωρίς εσώνια, ενώ τα γονίδια άλλων snRNA βρίσκονται μέσα στα ιντρόνια των γονιδίων που κωδικοποιούν πρωτεΐνες. Έτσι, στο Xenopus το U13 κωδικοποιείται από τρεις μοναδικές αλληλουχίες που βρίσκονται
τα εσώνια 5, 6 και 8 των γονιδίων της πρωτεΐνης θερμικού σοκ και το γονίδιο U16 βρίσκεται μέσα στο εσώνιο της ριβοσωμικής πρωτεΐνης L1. Η τελευταία περίσταση είναι σημαντική, αφού δείχνει ότι η επεξεργασία του rRNA και η επεξεργασία του mRNA των ριβοσωμικών πρωτεϊνών μπορεί να συντονιστεί με τη συμμετοχή του snRNA. Εκτός,
προτείνουν ότι τα snRNA είναι ικανά να χρησιμεύουν ως συνοδοί RNA, συμμετέχοντας στο αναδιπλούμενο RNA, δηλ. βοηθώντας την να δεχτεί την απαραίτητη δομή στο χώρο. Τα μικρά πυρηνικά RNA υπάρχουν στον πυρήνα σε σύμπλοκα με πρωτεΐνες που ονομάζονται μικρά σωματίδια ριβονουκλεοπρωτεΐνης (snRNPs). Ένα σταθερό συστατικό του snRNP είναι η πρωτεΐνη fibrillarin, μια πολύ δομικά συντηρητική πρωτεΐνη με μοριακό βάρος 34 kDa, εντοπισμένη στους πυρήνες. Ένα σύμπλεγμα που αποτελείται από πολλαπλά snRNPs που καταλύει το μάτισμα των πυρηνικών pro-mRNA ονομάζεται ματοσώματα .]

Είναι γνωστό ότι το snRNA του τύπου U 1 περιέχει μια νουκλεοτιδική αλληλουχία ομόλογη με το 5" άκρο του ιντρονίου. Το ζευγάρωμα αυτών των αλληλουχιών δημιουργεί ένα μάτισμα. Στη συνέχεια το snRNA των τύπων U2, U4, U5 και U6 ενώνει και αρχίζει το μάτισμα. Όπως και στην περίπτωση των εσωνίων της πρώτης ομάδας, δύο αντιδράσεις μετεστεροποίησης. Ζ"-η υδροξυλομάδα της αδενίνης (Α), που εντοπίζεται στο εσώνιο, αλληλεπιδρά με τη θέση ματίσματος 5", κόβοντας την αλυσίδα RNA. Στη συνέχεια, πολλά snRNP σχηματίζουν ένα ενδιάμεσο σύμπλεγμα και αρχίζει η δεύτερη αντίδραση: το ελεύθερο άκρο 5" του ιντρονίου συνδέεται με το υπόλειμμα αδενίνης. Ως αποτέλεσμα, σχηματίζεται μια χαρακτηριστική δομή τύπου λάσο που μοιάζει με βρόχο που περιέχει το διαγραμμένο εσώνιο. Στη συνέχεια, τα άκρα των εξονίων συνδέονται και το σύμπλεγμα snRNA απελευθερώνει το αντίγραφο .
[ Konichev, σ.294. Η αλληλεπίδραση των διαφόρων snRNA που αποτελούν μέρος του ματίσματος με το ματισμένο προ-mRNA στις θέσεις 5" και 3" δίνει στο εσώνιο μια δομή που μοιάζει με βρόχο. Σε αυτή την περίπτωση, τα άκρα των εξονίων έρχονται πιο κοντά μεταξύ τους, κάτι που διευκολύνεται από το σχηματισμό μη κανονικών (διαφορετικών από τα ζεύγη Watson-Crick) δεσμών υδρογόνου μεταξύ των δύο γουανινών που περιέχονται στις θέσεις ματίσματος 5" και 3" (βλ. Εικ. 148). Η συνένωση των εξονίων δημιουργεί μια προϋπόθεση για την επίθεση του άκρου 3" του ιντρονίου από ένα νουκλεοτίδιο αδενυλίου που βρίσκεται κοντά στο άκρο 3". Ως αποτέλεσμα της ρήξης του φωσφοδιεστερικού δεσμού μεταξύ του εξονίου 1 και του 5"-άκρου του ιντρονίου, το τελευταίο αλληλεπιδρά με το νουκλεοτίδιο αδενυλίου και το σχηματισμό ενός βρόχου τύπου λάσο στο εσώνιο (βλ. Εικ. 148_Konichev). αυτό, το απελευθερωμένο άκρο 3"-OH του εξονίου 1 κόβει τη θέση ματίσματος 3"-, διασπά το εσώνιο και, συνδέοντας με το εξόνιο 2, σχηματίζει τελικά ένα ώριμο (ματισμένο) μόριο mRNA ]
Επεξεργασία - αυτή είναι η ωρίμανση του preRNA που συντίθεται στο DNA και η μετατροπή του σε ώριμο RNA. Λαμβάνει χώρα στον πυρήνα του κυττάρου στους ευκαρυώτες.
Συστατικά της επεξεργασίας
- Μετακίνησηνουκλεοτίδια. Αποτέλεσμα: σημαντική μείωση στο μήκος και τη μάζα του αρχικού RNA.
- Ενταξηνουκλεοτίδια. Αποτέλεσμα: μια ελαφρά αύξηση στο μήκος και τη μάζα του αρχικού RNA.
- Τροποποίηση(τροποποίηση) νουκλεοτιδίων. Αποτέλεσμα: η εμφάνιση σπάνιων «εξωτικών» δευτερευόντων («μικρότερων») νουκλεοτιδίων στο RNA.
Αφαίρεση νουκλεοτιδίων
1. Χωρισμός μεμονωμένα νουκλεοτίδια, ένα κάθε φορά, από τα άκρα της αλυσίδας RNA. Εκτελείται από ένζυμα εξωνουκλεάσες. Τυπικά, το preRNA ξεκινά στο 5" άκρο του ATP ή του GTP και τελειώνει στο άκρο 3" με τις περιοχές GC. Χρειάζονται μόνο για την ίδια τη μεταγραφή, αλλά δεν χρειάζονται για να λειτουργήσει το RNA, επομένως διαχωρίζονται.
2. Αποκοπή Θραύσματα RNA που αποτελούνται από πολλά νουκλεοειδή. Εκτελείται από ένζυμα ενδονουκλεάσες. Με αυτόν τον τρόπο, οι αλληλουχίες νουκλεοτιδίων διαχωρισμού απομακρύνονται από τα άκρα του preRNA.
3. Τομή preRNA σε μεμονωμένα μόρια RNA. Εκτελείται από ένζυμα ενδονουκλεάσης. Με αυτόν τον τρόπο, λαμβάνεται ριβοσωμικό RNA (rRNA) και RNA ιστόνης (mRNA).
4. Συναρμολόγηση . Αυτό τομή μεσαίες τομές (ιντρονικές αλληλουχίες) από το preRNA και στη συνέχεια του ράψιμο . Η εκτομή πραγματοποιείται με ένζυμα ενδονουκλεάσης και η διασύνδεση πραγματοποιείται με λιγάσες. Το αποτέλεσμα είναι mRNA που αποτελείται μόνο από εξωνικές αλληλουχίες νουκλεοτιδίων. Όλα τα προ-mRNA είναι ματισμένα, εκτός από τα ιστονικά.
Ως αποτέλεσμα της αφαίρεσης νουκλεοτιδίων στο mRNA, για παράδειγμα, αντί για 9200 νουκλεοτίδια, μπορεί να παραμείνουν μόνο 1200.
Κατά μέσο όρο, μετά την επεξεργασία, μόνο το 13% του μήκους του προ-mRNA παραμένει στο ώριμο mRNA και το 87% χάνεται.
Προσθήκη νουκλεοτιδίων
Ένα τροποποιημένο νουκλεοτίδιο 7-μεθυλγουανυλίου συνδέεται στο pre-mRNA από το αρχικό άκρο 5" χρησιμοποιώντας έναν άτυπο πυροφωσφορικό δεσμό· αυτό είναι ένα συστατικό "καπάκι" ("καπέλα") mRNA. Αυτό το κάλυμμα δημιουργείται στο αρχικό στάδιο της σύνθεσης RNA για να προστατεύσει το εκκολαπτόμενο RNA από επιθέσεις των ενζύμων εξωνουκλεασών που αποκόπτουν τα τερματικά νουκλεοτίδια από το RNA.
Μετά την ολοκλήρωση της σύνθεσης του pre-mRNA, τα νουκλεοτίδια αδενυλίου προστίθενται διαδοχικά στο τελικό του τμήμα από το 3" άκρο από το ένζυμο πολυαδενυλική πολυμεράση, έτσι ώστε ένα πολυαδενυλικό "ουρά" περίπου 200-250 Α-νουκλεοτίδια. Οι στόχοι για αυτή τη διαδικασία είναι οι αλληλουχίες AAAAAAA και GGUUGUUGGUU στο τέλος του preRNA. Ως αποτέλεσμα, η ίδια η ουρά του preRNA κόβεται και αντικαθίσταται με μια ουρά polyA.
Βίντεο:Προμήθεια preRNA με κάλυμμα και ουρά
στο προ- tRNA ουρά στο 3" άκρο του δημιουργείται με τη διαδοχική προσθήκη τριών νουκλεοτιδίων: C, C και A. Αποτελούν τον κλάδο δέκτη του RNA μεταφοράς.
Τροποποίηση νουκλεοτιδίων
Είναι σημαντικό να σημειωθεί ότι τροποποιημένα δευτερεύοντα νουκλεοτίδια εμφανίζονται στο ώριμο RNA ως αποτέλεσμα της επεξεργασίας και δεν ενσωματώνονται στο RNA κατά τη σύνθεσή του στο DNA.
Στα νουκλεοτίδια του καλύμματος υπάρχουν mRNA η ριβόζη μεθυλιώνεται.
σε προ- rRNA Τα υπολείμματα ριβόζης μεθυλιώνονται επιλεκτικά σε όλο το μήκος της αλυσίδας, με συχνότητα περίπου 1%, δηλ. 1 νουκλεοτίδιο στα 100.
σε προ- tRNA η τροποποίηση γίνεται με τους πιο ποικίλους τρόπους. Για παράδειγμα, αν η ουριδίνη αναχθεί, γίνεται διυδροουριδίνη, αν ισομεριστεί, γίνεται ψευδουριδίνη, αν μεθυλιωθεί, γίνεται μεθυλουριδίνη Η αδενοσίνη μπορεί να απαμινωθεί και να μετατραπεί σε ινοσίνη και αν στη συνέχεια μεθυλιωθεί γίνεται μεθυλινοσίνη. Συμβαίνουν επίσης και άλλες τροποποιήσεις νουκλεοτιδίων.
Βίντεο:Λεπτομέρειες για την επεξεργασία
Αποτέλεσμα επεξεργασίας
Τα αρχικά preRNA συντομεύονται και τροποποιούνται . Τα κύτταρα εμφανίζονται στον πυρήνα ώριμο RNA ΔΙΑΦΟΡΕΤΙΚΟΙ ΤΥΠΟΙ: rRNA (28S, 18S, 5.8S, 5S), tRNA (1-3 τύποι για καθένα από τα 20 αμινοξέα), mRNA (χιλιάδες επιλογές ανάλογα με τον αριθμό των γονιδίων που εκφράζονται σε ένα δεδομένο κύτταρο). Εδώ στον πυρήνα, το rRNA συνδέεται με ριβοσωμικές πρωτεΐνες και σχηματίζει μεγάλες και μικρές ριβοσωμικές υπομονάδες. Φεύγουν από τον πυρήνα και εισέρχονται στο κυτταρόπλασμα. Και το mRNA δεσμεύεται για να μεταφέρει πρωτεΐνες και με αυτή τη μορφή εξέρχεται από τον πυρήνα στο κυτταρόπλασμα.
Τ ΤΕΡΜΑΤΙΣΜΟΣ
Η RNA πολυμεράση θα σταματήσει όταν φτάσει στα κωδικόνια τερματισμού. Με τη βοήθεια του παράγοντα τερματισμού πρωτεΐνης, του λεγόμενου παράγοντα ρ (ελληνικά ρ - «rho»), του ενζύμου και του μορίου RNA που συντίθεται. πρωταρχικό αντίγραφο, ο πρόδρομος του mRNA ή του tRNA ή του rRNA.
ΑΝΑΠΤΥΞΗ RNA
Αμέσως μετά τη σύνθεση, τα πρωτογενή μεταγραφήματα RNA, για διάφορους λόγους, δεν έχουν ακόμη δραστηριότητα, είναι «ανώριμα» και στη συνέχεια υφίστανται μια σειρά αλλαγών που ονομάζονται επεξεργασία. Στους ευκαρυώτες, όλοι οι τύποι προ-RNA υποβάλλονται σε επεξεργασία σε προκαρυωτικά, μόνο οι πρόδρομοι rRNA και tRNA.
ΕΠΕΞΕΡΓΑΣΙΑ ΤΟΥ ΠΡΟΔΡΟΜΟΥ MRNA
Κατά τη μεταγραφή τμημάτων DNA που μεταφέρουν πληροφορίες για πρωτεΐνες, σχηματίζονται ετερογενή πυρηνικά RNA, πολύ μεγαλύτερα σε μέγεθος από το mRNA. Το γεγονός είναι ότι λόγω της μωσαϊκής δομής των γονιδίων, αυτά τα ετερογενή RNA περιλαμβάνουν πληροφοριακά (εξόνια)
Και μη ενημερωτικό (ιντρόνια) περιοχές.
1. Το μάτισμα (eng. splice - to glue end to end) είναι μια ειδική διαδικασία κατά την οποία, με τη συμμετοχή μικρών πυρηνικών RNA, αφαιρούνται εσώνια και διατηρούνται τα εξόνια.

2. Capping (Αγγλικό καπάκι - καπέλο) - εμφανίζεται κατά τη μεταγραφή. Η διαδικασία συνίσταται στην προσθήκη του άνθρακα N7-μεθυλ-γουανοσίνης 5" στο 5"-τριφωσφορικό του τερματικού νουκλεοτιδίου του pre-mRNA.
Το «κάλυμμα» είναι απαραίτητο για την προστασία του μορίου RNA από τις εξωνουκλεάσες που λειτουργούν από το 5" άκρο, καθώς και για τη σύνδεση του mRNA στο ριβόσωμα και για την έναρξη της μετάφρασης.
3. Πολυαδενυλίωση– με τη βοήθεια πολυαδενυλικής πολυμεράσης χρησιμοποιώντας μόρια ATP, από 100 έως 200 νουκλεοτίδια αδενυλίου συνδέονται στο άκρο 3" του RNA, σχηματίζοντας μια ουρά πολυ(Α). Η ουρά πολυ(Α) είναι απαραίτητη για την προστασία του μορίου RNA από τις εξωνουκλεάσες δουλεύοντας με 3 "-τέλος.
ΕΠΕΞΕΡΓΑΣΙΑ ΤΟΥ ΠΡΟΔΡΟΜΟΥ RRNA
Οι πρόδρομοι rRNA είναι μεγαλύτερα μόρια σε σύγκριση με τα ώριμα rRNA. Η ωρίμανση τους καταλήγει στην κοπή του προριβοσωμικού RNA σε μικρότερες μορφές, οι οποίες εμπλέκονται άμεσα στο σχηματισμό του ριβοσώματος. Οι ευκαρυώτες έχουν 5S, 5.8S, 18S και 28S rRNA. Σε αυτή την περίπτωση, το 5S rRNA συντίθεται χωριστά και το μεγάλο προριβοσωμικό 45S RNA διασπάται από συγκεκριμένες νουκλεάσες για να σχηματιστεί
5.8S rRNA, 18S rRNA και 28S rRNA.
U Στους προκαρυώτες, τα μόρια του ριβοσωμικού RNA έχουν εντελώς διαφορετικές ιδιότητες(5S-, 16S-
23S-rRNA), το οποίο αποτελεί τη βάση για την εφεύρεση και τη χρήση ενός αριθμού αντιβιοτικών στην ιατρική
Π ROCESSING PRECEDOR T RNA
1. Σχηματισμός στο άκρο 3" της αλληλουχίας C-C-A. Για αυτό, κάποιοιπρο-tRNA από το άκρο 3". Η περίσσεια νουκλεοτιδίων αφαιρείται μέχρι να «εκτεθεί» η τριάδαΓ-Γ-Α, για άλλους, αυτή η ακολουθία προστίθεται.
2. Σχηματισμός βρόχου αντικωδικονίουσυμβαίνει με μάτισμα και αφαίρεση ενός εσωνίου στο μεσαίο τμήμα του pre-tRNA.
3. Τροποποίηση νουκλεοτιδίωνστο μόριο με απαμίνωση, μεθυλίωση, αναγωγή. Για παράδειγμα, ο σχηματισμός ψευδουριδίνης και διυδροουριδίνης.
Κάτω από Η επεξεργασία του RNA κατανοεί τη διαδικασία ωρίμανσης του, που εμφανίζεται κατά τη διάρκεια και μετά τη μεταγραφή του και προηγείται της διαδικασίας μετάφρασης.
Διαφορετικοί τύποι RNA επεξεργάζονται διαφορετικά. Ωστόσο, στους προκαρυώτες, το αγγελιοφόρο RNA (mRNA) δεν υποβάλλεται σε επεξεργασία. Τυπικά, η επεξεργασία RNA θεωρείται χρησιμοποιώντας ευκαρυωτικό mRNA ως παράδειγμα.
Όπως γνωρίζετε, το RNA συντίθεται σε ένα τμήμα μιας από τις αλυσίδες του DNA και αυτή η διαδικασία ονομάζεται μεταγραφή. Στα σχολικά μαθήματα, η μεταγραφή συνήθως ακολουθείται αμέσως από τη διαδικασία της μετάφρασης, στην οποία το mRNA χρησιμοποιείται ως πρότυπο για τη σύνθεση πρωτεϊνών. Ωστόσο, μεταξύ της μεταγραφής και της μετάφρασης, συμβαίνει ένας αριθμός μετασχηματισμών με το RNA, με αποτέλεσμα να γίνεται λειτουργικά ενεργό. Αυτές οι τροποποιήσεις ονομάζονται συλλογικά επεξεργασία. Ορισμένα από τα στάδιά του εμφανίζονται ήδη κατά τη στιγμή της μεταγραφής.
Ας εξετάσουμε την επεξεργασία του ευκαρυωτικού αγγελιοφόρου (αγγελιοφόρου) RNA.
Κάλυψη. Ακόμη και στο στάδιο της μεταγραφής, ένα μόριο μεθυλγουανοσίνης, που είναι μια μεθυλιωμένη αζωτούχα βάση γουανοσίνη, συνδέεται στο αρχικό (5") άκρο του μορίου RNA μέσω μιας γέφυρας τριφωσφορικού (τρία υπολείμματα φωσφορικού οξέος). Επίσης, τα υπολείμματα ριβόζης μεθυλιώνονται σε τα δύο πρώτα νουκλεοτίδια του mRNA Αυτές οι διεργασίες ονομάζονται κάλυψη, και ο σχηματισμός του καπάκι(καπέλο). Προστατεύει το μόριο από την ενζυματική αποικοδόμηση, συμμετέχει σε άλλα στάδια επεξεργασίας και ξεκινά τη μετάφραση.
Πολυαδενυλίωση. Μετά την ολοκλήρωση της μεταγραφής, πολλά νουκλεοτίδια αδενίνης (από 100 έως 250) συνδέονται στο άκρο (3") του RNA. Σχηματίζεται ένα πολυαδενυλικό άκρο - πολυ-Α. Επιτελεί επίσης προστατευτική λειτουργία, αποτρέποντας τη δράση καταστροφικών ενζύμων.
Συναρμολόγηση. Ένα πρόδρομο μόριο mRNA (προ-mRNA) είναι ένα αντίγραφο ενός τμήματος DNA (γονίδιο) που περιλαμβάνει αμετάφραστες περιοχές (που βρίσκονται στα άκρα) και εναλλασσόμενα εσώνια και εξόνια. Τα εσώνια δεν συμμετέχουν στη μετάφραση και πρέπει να αφαιρεθούν πριν από αυτήν. Το μάτισμα είναι η διαδικασία κοπής του mRNA, αφαίρεσης εσωνίων και συρραφής των υπόλοιπων εξονίων.
Ως αποτέλεσμα του ματίσματος, το μήκος του μορίου mRNA μειώνεται σημαντικά. Η διαδικασία καταλύεται από ένα ειδικό σύμπλεγμα - μάτισμα, συμπεριλαμβανομένων μικρών πυρηνικών RNA και ενζυμικών πρωτεϊνών. Τα εξόνια μπορούν να συρράπτονται μεταξύ τους με διαφορετικούς τρόπους (εναλλάξ με διαφορετικούς τρόπους, μερικά μπορούν να παραλειφθούν). Αυτό το φαινόμενο ονομάζεται εναλλακτικό μάτισμα. Ως αποτέλεσμα, ένα προ-mRNA μπορεί να παράγει πολλά διαφορετικά mRNA, στα οποία θα συντεθούν διαφορετικές πρωτεΐνες.
Τα RNA μεταφοράς (tRNA) επίσης συχνά υποβάλλονται σε επεξεργασία. Ωστόσο, το δικό τους είναι διαφορετικό και σχετίζεται κυρίως με τη μεθυλίωση μεμονωμένων νουκλεοτιδίων. Ως αποτέλεσμα, το tRNA παίρνει τη χαρακτηριστική του μορφή και γίνεται ενεργό (ικανό να δεσμεύεται με αμινοξέα).
Η επεξεργασία του ριβοσωμικού RNA (rRNA) περιορίζεται κυρίως στην αποκοπή του κοινού μεταγράφου (προ-rRNA), από το οποίο μέρη σχηματίζουν τρία διαφορετικά μόρια rRNA (από τα τέσσερα).
Μετά την επεξεργασία, ώριμα μόρια mRNA, tRNA και σχηματισμένα ριβοσωματικά υποσωματίδια (που περιέχουν rRNA) μεταφέρονται από τον πυρήνα στο κυτταρόπλασμα, όπου, εκτελώντας κάθε ρόλο τους, εξασφαλίζουν τη διαδικασία μετάφρασης (σύνθεση πρωτεϊνών).
Αυτό το στάδιο είναι που διακρίνει την εφαρμογή της υπάρχουσας γενετικής πληροφορίας σε κύτταρα όπως οι ευκαρυώτες και οι προκαρυώτες.
Ερμηνεία αυτής της έννοιας
Μεταφρασμένος από τα αγγλικά, αυτός ο όρος σημαίνει «επεξεργασία, επεξεργασία». Η επεξεργασία είναι η διαδικασία παραγωγής ώριμων μορίων ριβονουκλεϊκού οξέος από προ-RNA. Με άλλα λόγια, πρόκειται για ένα σύνολο αντιδράσεων που οδηγούν στον μετασχηματισμό των πρωτογενών προϊόντων μεταγραφής (προ-RNA διαφόρων τύπων) σε ήδη λειτουργικά μόρια.
Όσο για την επεξεργασία του r- και του tRNA, συνήθως καταλήγει στην αποκοπή περιττών θραυσμάτων από τα άκρα των μορίων. Αν μιλάμε για mRNA, τότε μπορεί να σημειωθεί ότι στους ευκαρυώτες αυτή η διαδικασία συμβαίνει με τρόπο πολλαπλών σταδίων.
Έτσι, αφού έχουμε ήδη μάθει ότι η επεξεργασία είναι ο μετασχηματισμός ενός πρωτογενούς μεταγράφου σε ένα ώριμο μόριο RNA, αξίζει να προχωρήσουμε στην εξέταση των χαρακτηριστικών του.
Κύρια χαρακτηριστικά της υπό εξέταση έννοιας
Αυτά περιλαμβάνουν τα ακόλουθα:
- τροποποίηση και των δύο άκρων του μορίου και του RNA, κατά την οποία προσαρτώνται συγκεκριμένες αλληλουχίες νουκλεοτιδίων, υποδεικνύοντας τη θέση της αρχής (τέλους) της μετάφρασης.
- Το μάτισμα είναι η αποκοπή μη πληροφοριακών αλληλουχιών ριβονουκλεϊκών οξέων που αντιστοιχούν σε ιντρόνια DNA.
Όσον αφορά τα προκαρυωτικά, το mRNA τους δεν υπόκειται σε επεξεργασία. Έχει την ικανότητα να λειτουργεί αμέσως μετά την ολοκλήρωση της σύνθεσης.
Πού λαμβάνει χώρα η εν λόγω διαδικασία;
Σε κάθε οργανισμό, η επεξεργασία του RNA λαμβάνει χώρα στον πυρήνα. Διεξάγεται μέσω ειδικών ενζύμων (μια ομάδα από αυτά) για κάθε μεμονωμένο τύπο μορίου. Τα προϊόντα μετάφρασης όπως τα πολυπεπτίδια που διαβάζονται απευθείας από το mRNA μπορούν επίσης να υποβληθούν σε επεξεργασία. Τα λεγόμενα πρόδρομα μόρια των περισσότερων πρωτεϊνών - κολλαγόνο, ανοσοσφαιρίνες, πεπτικά ένζυμα, ορισμένες ορμόνες - υφίστανται αυτές τις αλλαγές, μετά τις οποίες αρχίζει η πραγματική τους λειτουργία στο σώμα.
Έχουμε ήδη μάθει ότι η επεξεργασία είναι η διαδικασία σχηματισμού ώριμου RNA από προ-RNA. Τώρα αξίζει να εμβαθύνουμε στη φύση του ίδιου του ριβονουκλεϊκού οξέος.
RNA: χημική φύση
Είναι ένα συμπολυμερές ριβονουκλεοτιδίων πυριμιδίνης και πουρίνης, τα οποία συνδέονται μεταξύ τους, όπως και στο DNA, με φωσφοδιεστερικές γέφυρες 3' - 5'.

Αν και αυτοί οι δύο τύποι μορίων είναι παρόμοιοι, διαφέρουν με διάφορους τρόπους.
Διακριτικά χαρακτηριστικά του RNA και του DNA
Πρώτον, το ριβονουκλεϊκό οξύ έχει ένα υπόλειμμα άνθρακα, το οποίο βρίσκεται δίπλα σε βάσεις πυριμιδίνης και πουρίνης, φωσφορικές ομάδες - ριβόζη, ενώ το DNA έχει 2'-δεοξυριβόζη.
Δεύτερον, τα συστατικά πυριμιδίνης είναι επίσης διαφορετικά. Παρόμοια συστατικά είναι τα νουκλεοτίδια της αδενίνης, της κυτοσίνης και της γουανίνης. Το RNA περιέχει ουρακίλη αντί για θυμίνη.

Τρίτον, το RNA έχει μια 1-κλωνη δομή και το DNA είναι ένα 2-κλώνο μόριο. Αλλά στην αλυσίδα του ριβονουκλεϊκού οξέος υπάρχουν περιοχές με αντίθετη πολικότητα (συμπληρωματική αλληλουχία), χάρη στις οποίες η μονή αλυσίδα του μπορεί να διπλώσει και να σχηματίσει "φουρκέτες" - δομές προικισμένες με 2-έλικα χαρακτηριστικά (όπως φαίνεται στο παραπάνω σχήμα).
Τέταρτον, λόγω του γεγονότος ότι το RNA είναι μια μονή αλυσίδα που είναι συμπληρωματική μόνο σε μία από τις αλυσίδες DNA, η γουανίνη δεν χρειάζεται απαραίτητα να υπάρχει σε αυτό με το ίδιο περιεχόμενο με την κυτοσίνη και την αδενίνη - όπως η ουρακίλη.
Πέμπτον, το RNA μπορεί να υδρολυθεί με αλκάλια σε 2', 3'-κυκλικούς διεστέρες μονονουκλεοτιδίων. Το ρόλο του ενδιάμεσου προϊόντος στην υδρόλυση παίζει ο 2', 3', 5-τριεστέρας, ο οποίος είναι ανίκανος να σχηματιστεί για DNA κατά τη διάρκεια μιας παρόμοιας διαδικασίας λόγω της απουσίας 2'-υδροξυλικών ομάδων. Σε σύγκριση με το DNA, η αλκαλική αστάθεια του ριβονουκλεϊκού οξέος είναι μια χρήσιμη ιδιότητα τόσο για διαγνωστικούς όσο και για αναλυτικούς σκοπούς.

Αυτή η αλληλουχία είναι συμπληρωματική της γονιδιακής αλυσίδας (που κωδικοποιεί) από την οποία «διαβάζεται» το RNA. Λόγω αυτής της ιδιότητας, το μόριο του ριβονουκλεϊκού οξέος μπορεί να συνδεθεί ειδικά με τον κωδικοποιητικό κλώνο, αλλά δεν μπορεί να το κάνει αυτό με τον μη κωδικοποιητικό κλώνο DNA. Η αλληλουχία RNA, εκτός από την αντικατάσταση του Τ με U, είναι παρόμοια με αυτή του μη κωδικοποιητικού κλώνου του γονιδίου.
Τύποι RNA
Σχεδόν όλοι εμπλέκονται σε μια τέτοια διαδικασία όπως είναι γνωστοί οι ακόλουθοι τύποι RNA:
- Πρότυπο (mRNA).Αυτά είναι μόρια κυτταροπλασματικού ριβονουκλεϊκού οξέος που χρησιμεύουν ως πρότυπα για τη σύνθεση πρωτεϊνών.
- Ριβοσωμικό (rRNA).Αυτό είναι ένα μόριο κυτταροπλασμικού RNA που παίζει το ρόλο τέτοιων δομικών συστατικών όπως τα ριβοσώματα (οργανίδια που εμπλέκονται στη σύνθεση πρωτεϊνών).
- Μεταφορά (tRNA). Αυτά είναι μόρια που συμμετέχουν στη μετάφραση (μετάφραση) των πληροφοριών mRNA σε μια αλληλουχία αμινοξέων ήδη σε πρωτεΐνες.
Ένα σημαντικό μέρος του RNA με τη μορφή πρώτων μεταγραφών, που σχηματίζονται και σε κύτταρα θηλαστικών, υπόκειται σε αποικοδόμηση στον πυρήνα και δεν παίζει πληροφοριακό ή δομικό ρόλο στο κυτταρόπλασμα.
Σε ανθρώπινα κύτταρα (καλλιεργημένα) έχει βρεθεί μια κατηγορία μικρών πυρηνικών ριβονουκλεϊκών οξέων που δεν εμπλέκονται άμεσα στη σύνθεση πρωτεϊνών, αλλά έχουν αντίκτυπο στην επεξεργασία του RNA, καθώς και στη γενική κυτταρική «αρχιτεκτονική». Τα μεγέθη τους ποικίλλουν, περιέχουν 90 - 300 νουκλεοτίδια.
Το ριβονουκλεϊκό οξύ είναι το κύριο γενετικό υλικό ενός αριθμού φυτικών και ζωικών ιών. Μερικοί ιοί RNA δεν περνούν ποτέ από το στάδιο RNA στο DNA. Ωστόσο, πολλοί ζωικοί ιοί, για παράδειγμα ρετροϊοί, χαρακτηρίζονται από αντίστροφη μετάφραση του γονιδιώματος RNA τους, κατευθυνόμενη από εξαρτώμενη από RNA ανάστροφη μεταγραφάση (DNA πολυμεράση) με το σχηματισμό αντιγράφου 2-κλωνου DNA. Στις περισσότερες περιπτώσεις, το αναδυόμενο μεταγράφημα 2-κλώνου DNA ενσωματώνεται στο γονιδίωμα, διασφαλίζοντας στη συνέχεια την έκφραση ιικών γονιδίων και την παραγωγή νέων αντιγράφων γονιδιωμάτων RNA (επίσης ιικού).
Μετα-μεταγραφικές τροποποιήσεις του ριβονουκλεϊκού οξέος
Τα μόριά του, που συντίθενται με RNA πολυμεράσες, είναι πάντα λειτουργικά ανενεργά και δρουν ως πρόδρομοι, δηλαδή προ-RNA. Μετατρέπονται σε ήδη ώριμα μόρια μόνο αφού υποστούν τις αντίστοιχες μετα-μεταγραφικές τροποποιήσεις του RNA - τα στάδια της ωρίμανσης του.
Ο σχηματισμός των ώριμων mRNAs ξεκινά κατά τη διάρκεια της σύνθεσης RNA και της πολυμεράσης II στο στάδιο της επιμήκυνσης. Ήδη στο 5'-άκρο του σταδιακά αναπτυσσόμενου κλώνου RNA, το GTP συνδέεται με το 5'-άκρο και στη συνέχεια το ορθοφωσφορικό αποκόπτεται. Η γουανίνη στη συνέχεια μεθυλιώνεται για να σχηματίσει 7-μεθυλ-GTP. Αυτή η ειδική ομάδα που βρίσκεται στο mRNA ονομάζεται «καπάκι» (cap).
Ανάλογα με τον τύπο του RNA (ριβοσωμικό, μεταφορά, εκμαγείο κ.λπ.), οι πρόδρομοι υφίστανται διάφορες διαδοχικές τροποποιήσεις. Για παράδειγμα, οι πρόδρομοι mRNA υφίστανται μάτισμα, μεθυλίωση, κάλυψη, πολυαδενυλίωση και μερικές φορές επεξεργασία.

Ευκαρυώτες: γενικά χαρακτηριστικά
Το ευκαρυωτικό κύτταρο δρα ως περιοχή ζωντανών οργανισμών και περιέχει τον πυρήνα. Εκτός από τα βακτήρια, τα αρχαία, όλοι οι οργανισμοί είναι πυρηνικοί. Τα φυτά, οι μύκητες, τα ζώα, συμπεριλαμβανομένης μιας ομάδας οργανισμών που ονομάζονται πρωτίστες, είναι όλοι ευκαρυωτικοί οργανισμοί. Μπορούν να είναι τόσο μονοκύτταρα όσο και πολυκύτταρα, αλλά όλα έχουν ένα κοινό σχέδιο κυτταρικής δομής. Είναι γενικά αποδεκτό ότι αυτοί οι πολύ διαφορετικοί οργανισμοί έχουν την ίδια προέλευση, γι' αυτό η πυρηνική ομάδα γίνεται αντιληπτή ως μονοφυλετική ταξινόμηση της υψηλότερης τάξης.
Με βάση κοινές υποθέσεις, οι ευκαρυώτες εμφανίστηκαν πριν από 1,5 - 2 δισεκατομμύρια χρόνια. Σημαντικός ρόλος στην εξέλιξή τους δίνεται στη συμβιογένεση -τη συμβίωση ενός ευκαρυωτικού κυττάρου που είχε έναν πυρήνα ικανό για φαγοκυττάρωση και βακτήρια που καταπίνονται από αυτόν - οι πρόδρομοι των πλαστιδίων και των μιτοχονδρίων.
Προκαρυώτες: γενικά χαρακτηριστικά
Πρόκειται για ζωντανούς οργανισμούς ενός κυττάρου που δεν έχουν πυρήνα (σχηματισμένο) και άλλα μεμβρανικά οργανίδια (εσωτερικά). Το μόνο μεγάλο κυκλικό 2κλωνο μόριο DNA που περιέχει τον κύριο όγκο του γενετικού υλικού ενός κυττάρου είναι αυτό που δεν σχηματίζει σύμπλοκο με πρωτεΐνες ιστόνης.
Τα προκαρυωτικά περιλαμβάνουν αρχαία και βακτήρια, συμπεριλαμβανομένων των κυανοβακτηρίων. Οι απόγονοι των κυττάρων χωρίς πυρήνα είναι ευκαρυωτικά οργανίδια - πλαστίδια, μιτοχόνδρια. Χωρίζονται σε 2 taxa εντός της κατάταξης τομέα: Archaea και Bacteria.
Αυτά τα κύτταρα δεν έχουν πυρηνική μεμβράνη, η συσκευασία του DNA συμβαίνει χωρίς τη συμμετοχή ιστονών. Ο τύπος τροφοδοσίας τους είναι οσμοτροφικός και το γενετικό υλικό αντιπροσωπεύεται από ένα που είναι κλειστό σε έναν δακτύλιο και υπάρχει μόνο 1 ρεπλικόνιο. Οι προκαρυώτες εξακολουθούν να έχουν οργανίδια που έχουν δομή μεμβράνης.
Διαφορά μεταξύ ευκαρυωτών και προκαρυωτών
Το θεμελιώδες χαρακτηριστικό των ευκαρυωτικών κυττάρων συνδέεται με την παρουσία της γενετικής συσκευής σε αυτά, η οποία βρίσκεται στον πυρήνα, όπου προστατεύεται από μια μεμβράνη. Το DNA τους είναι γραμμικό, σχετίζεται με πρωτεΐνες ιστόνης, άλλες χρωμοσωμικές πρωτεΐνες που απουσιάζουν στα βακτήρια. Κατά κανόνα περιέχουν 2 πυρηνικές φάσεις. Το ένα έχει ένα απλοειδές σύνολο χρωμοσωμάτων, και στη συνέχεια συγχωνεύονται, 2 απλοειδή κύτταρα σχηματίζουν ένα διπλοειδές, το οποίο περιέχει ήδη ένα 2ο σύνολο χρωμοσωμάτων. Συμβαίνει επίσης ότι με επακόλουθη διαίρεση το κύτταρο γίνεται ξανά απλοειδές. Αυτό το είδος του κύκλου ζωής, καθώς και η διπλοειδικότητα γενικά, δεν είναι τυπικό για τους προκαρυώτες.
Η πιο ενδιαφέρουσα διαφορά είναι η παρουσία ειδικών οργανιδίων στους ευκαρυώτες, τα οποία έχουν τη δική τους γενετική συσκευή και αναπαράγονται με διαίρεση. Αυτές οι δομές περιβάλλονται από μια μεμβράνη. Αυτά τα οργανίδια είναι πλαστίδια και μιτοχόνδρια. Όσον αφορά τη δραστηριότητα και τη δομή της ζωής τους, είναι εκπληκτικά παρόμοια με τα βακτήρια. Αυτή η περίσταση ώθησε τους επιστήμονες να σκεφτούν ότι είναι απόγονοι βακτηριακών οργανισμών που μπήκαν σε συμβίωση με ευκαρυώτες.
Τα προκαρυωτικά έχουν μικρό αριθμό οργανιδίων, κανένα από τα οποία δεν περιβάλλεται από δεύτερη μεμβράνη. Δεν έχουν ενδοπλασματικό δίκτυο και λυσοσώματα.
Μια άλλη σημαντική διαφορά μεταξύ των ευκαρυωτών και των προκαρυωτικών είναι η παρουσία του φαινομένου της ενδοκυττάρωσης στους ευκαρυώτες, συμπεριλαμβανομένης της φαγοκυττάρωσης στις περισσότερες ομάδες. Το τελευταίο είναι η ικανότητα σύλληψης, εγκλεισμού σε ένα κυστίδιο μεμβράνης, και στη συνέχεια πέψης διαφόρων στερεών σωματιδίων. Αυτή η διαδικασία παρέχει την πιο σημαντική προστατευτική λειτουργία στο σώμα. Η εμφάνιση φαγοκυττάρωσης οφείλεται πιθανώς στο γεγονός ότι τα κύτταρα τους είναι μεσαίου μεγέθους. Οι προκαρυωτικοί οργανισμοί είναι δυσανάλογα μικρότεροι, γι' αυτό και κατά την εξέλιξη των ευκαρυωτών προέκυψε η ανάγκη που σχετίζεται με τον εφοδιασμό του κυττάρου με σημαντική ποσότητα τροφής. Ως αποτέλεσμα, ανάμεσά τους εμφανίστηκαν τα πρώτα κινητά αρπακτικά.

Η επεξεργασία ως ένα από τα στάδια της βιοσύνθεσης πρωτεϊνών
Αυτό είναι το δεύτερο στάδιο, το οποίο ξεκινά μετά τη μεταγραφή. Η επεξεργασία πρωτεϊνών συμβαίνει μόνο σε ευκαρυώτες. Αυτή είναι η ωρίμανση του mRNA. Για την ακρίβεια, πρόκειται για την αφαίρεση περιοχών που δεν κωδικοποιούν μια πρωτεΐνη και την προσθήκη μαρτύρων.

συμπέρασμα
Αυτό το άρθρο περιγράφει τι είναι η επεξεργασία (βιολογία). Εξηγεί επίσης τι είναι το RNA, παρατίθενται οι τύποι του και οι μετα-μεταγραφικές τροποποιήσεις. Εξετάζονται τα διακριτικά χαρακτηριστικά των ευκαρυωτών και των προκαρυωτικών.
Τέλος, αξίζει να υπενθυμίσουμε ότι η επεξεργασία είναι η διαδικασία σχηματισμού ώριμου RNA από προ-RNA.
- Σε επαφή με 0
- Google+ 0
- Εντάξει 0
- Facebook 0