Χρησιμοποιώντας ένα ηλεκτρονικό μικροσκόπιο, μπορεί να φανεί ένα ινιδιακό δίκτυο στο κυτταρόπλασμα των ευκαρυωτικών, οι λειτουργίες του οποίου σχετίζονται με την κίνηση του ενδοκυτταρικού περιεχομένου, την κίνηση του ίδιου του κυττάρου και επίσης, σε συνδυασμό με τη διατήρηση του σχήματος του κυττάρου. Ένα από αυτά τα ινίδια είναι μικροσωληνίσκους(συνήθως από μερικά μικρόμετρα έως μερικά χιλιοστά σε μήκος), που αντιπροσωπεύουν μακριούς λεπτούς κυλίνδρους(διάμετρος περίπου 25 nm) με μια κοιλότητα μέσα.Ταξινομούνται ως κυτταρικά οργανίδια.
Τα τοιχώματα των μικροσωληνίσκων αποτελούνται από ελικοειδή υπομονάδες πρωτεΐνης τουμπουλίνη, που αποτελείται από δύο μέρη, δηλαδή αντιπροσωπεύει ένα διμερές.
Οι παρακείμενοι σωλήνες μπορούν να συνδεθούν μεταξύ τους με προεξοχές των τοίχων τους.
Αυτό το κυτταρικό οργανίδιο είναι μια δυναμική δομή, έτσι μπορεί να αναπτυχθεί και να αποσυντεθεί (πολυμερίζεται και αποπολυμερίζεται). Η ανάπτυξη συμβαίνει λόγω της προσθήκης νέων υπομονάδων τουμπουλίνης στο ένα άκρο (συν) και της καταστροφής στο άλλο (μείον άκρο). Δηλαδή, οι μικροσωληνίσκοι είναι πολικοί.
Στα ζωικά κύτταρα (όπως και σε πολλά πρωτόζωα), τα κέντρα οργάνωσης των μικροσωληνίσκων είναι κεντριόλια. Οι ίδιοι αποτελούνται από εννέα τρίδυμα βραχυκυκλωμένων μικροσωληνίσκων και βρίσκονται κοντά στον πυρήνα. Από τα κεντρόλια, οι σωλήνες αποκλίνουν ακτινικά, αναπτύσσονται δηλαδή προς την περιφέρεια του κυττάρου. Στα φυτά, άλλες δομές λειτουργούν ως κέντρα οργάνωσης.
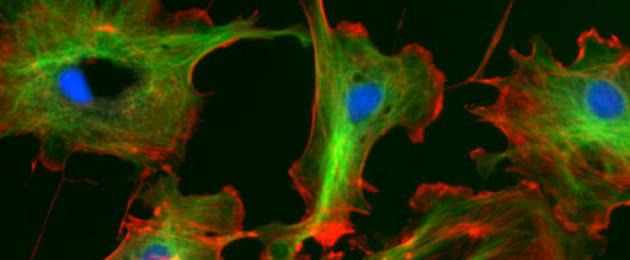
 Φθορισμός κυτταρικών ινιδίων (οι μικροσωληνίσκοι ανάβουν πράσινο)
Φθορισμός κυτταρικών ινιδίων (οι μικροσωληνίσκοι ανάβουν πράσινο) Η άτρακτος αποτελείται από μικροσωληνίσκους, οι οποίοι διαχωρίζουν τις χρωματίδες ή τα χρωμοσώματα κατά τη διάρκεια της μίτωσης ή της μείωσης. Αποτελούν τα βασικά σώματα που βρίσκονται στη βάση των βλεφαρίδων και των μαστιγίων. Η κίνηση της ατράκτου, των βλεφαρίδων και των μαστιγίων συμβαίνει λόγω της ολίσθησης των σωλήνων.

Μια παρόμοια λειτουργία είναι η κίνηση ενός αριθμού κυτταρικών οργανιδίων και σωματιδίων (για παράδειγμα, εκκριτικά κυστίδια που σχηματίζονται στη συσκευή Golgi, λυσοσώματα, ακόμη και μιτοχόνδρια). Σε αυτή την περίπτωση, οι μικροσωληνίσκοι παίζουν το ρόλο ενός είδους σιδηροτροχιών. Ειδικές πρωτεΐνες κινητήρα συνδέονται με τους σωλήνες στο ένα άκρο και στα οργανίδια στο άλλο. Λόγω της κίνησής τους κατά μήκος των σωλήνων, τα οργανίδια μεταφέρονται. Σε αυτή την περίπτωση, ορισμένες κινητικές πρωτεΐνες μετακινούνται μόνο από το κέντρο προς την περιφέρεια ( κινεσίνες), άλλα ( dyneins) - από την περιφέρεια προς το κέντρο.


Οι μικροσωληνίσκοι, λόγω της ακαμψίας τους, συμμετέχουν στο σχηματισμό του συστήματος υποστήριξης κυττάρων - κυτταροσκελετός. Προσδιορίστε το σχήμα του κελιού.
Η συναρμολόγηση και η αποσυναρμολόγηση των μικροσωληνίσκων, καθώς και η μεταφορά τους κατά μήκος αυτών, απαιτεί ενέργεια.
| Όνομα παραμέτρου | Εννοια |
| Θέμα άρθρου: | Μικροσωληνίσκοι |
| Ρουμπρίκα (θεματική κατηγορία) | Οικολογία |
Γενικά χαρακτηριστικά μικροσωληνίσκων.Τα υποχρεωτικά συστατικά του κυτταροσκελετού περιλαμβάνουν μικροσωληνίσκους (Εικ. 265), νηματώδεις μη διακλαδιζόμενες δομές, πάχους 25 nm, που αποτελούνται από πρωτεΐνες τουμπουλίνης και πρωτεΐνες που σχετίζονται με αυτές. Όταν πολυμερίζονται, οι τουμπουλίνες σχηματίζουν κοίλους σωλήνες (μικροσωληνίσκους), το μήκος των οποίων μπορεί να φτάσει αρκετά μικρά και οι μακρύτεροι μικροσωληνίσκοι βρίσκονται στο αξονικό τμήμα των ουρών του σπέρματος.
Οι μικροσωληνίσκοι βρίσκονται στο κυτταρόπλασμα των μεσοφασικών κυττάρων μεμονωμένα, σε μικρές χαλαρές δέσμες ή με τη μορφή πυκνά συσσωρευμένων σχηματισμών μέσα σε κεντρόλες, βασικά σώματα σε βλεφαρίδες και μαστίγια. Κατά τη διάρκεια της κυτταρικής διαίρεσης, οι περισσότεροι μικροσωληνίσκοι του κυττάρου αποτελούν μέρος της ατράκτου διαίρεσης.
Όσον αφορά τη δομή, οι μικροσωληνίσκοι είναι μακροί κοίλοι κύλινδροι με εξωτερική διάμετρο 25 nm (Εικ. 266). Το τοίχωμα του μικροσωληνίσκου αποτελείται από πολυμερισμένα μόρια πρωτεΐνης τουμπουλίνης. Κατά τον πολυμερισμό, τα μόρια της τουμπουλίνης σχηματίζουν 13 διαμήκεις πρωτονήματα, τα οποία καμπυλώνουν σε έναν κοίλο σωλήνα (Εικ. 267). Το μέγεθος του μονομερούς τουμπουλίνης είναι περίπου 5 nm, ίσο με το πάχος του τοιχώματος του μικροσωληνίσκου, στη διατομή του οποίου είναι ορατά 13 σφαιρικά μόρια.
Το μόριο της τουμπουλίνης είναι ένα ετεροδιμερές που αποτελείται από δύο διαφορετικές υπομονάδες, την α-τουμπουλίνη και τη β-τουμπουλίνη, οι οποίες, κατά τη σύνδεση, σχηματίζουν την ίδια την πρωτεΐνη τουμπουλίνης, η οποία είναι αρχικά πολωμένη. Και οι δύο υπομονάδες του μονομερούς τουμπουλίνης συνδέονται με το GTP, ωστόσο, στην α-υπομονάδα, το GTP δεν υφίσταται υδρόλυση, σε αντίθεση με το GTP στην β-υπομονάδα, όπου, κατά τον πολυμερισμό, λαμβάνει χώρα υδρόλυση του GTP στο GDP. Κατά τον πολυμερισμό, τα μόρια τουμπουλίνης συνδυάζονται με τέτοιο τρόπο ώστε η α-υπομονάδα της επόμενης πρωτεΐνης να συνδέεται με την β-υπομονάδα μιας πρωτεΐνης κ.λπ. Κατά συνέπεια, μεμονωμένα πρωτοϊνίδια προκύπτουν ως πολικά νημάτια, και συνεπώς ολόκληρος ο μικροσωληνίσκος είναι επίσης μια πολική δομή, που έχει ένα ταχέως αναπτυσσόμενο (+) άκρο και ένα αργά αναπτυσσόμενο (-) άκρο (Εικ. 268).
Όταν η συγκέντρωση πρωτεΐνης είναι επαρκής, ο πολυμερισμός συμβαίνει αυθόρμητα. Αλλά κατά τη διάρκεια του αυθόρμητου πολυμερισμού των τουμπουλινών, λαμβάνει χώρα υδρόλυση ενός μορίου GTP που σχετίζεται με τη β-τουμπουλίνη. Κατά την επιμήκυνση του μικροσωληνίσκου, η δέσμευση της τουμπουλίνης εμφανίζεται με υψηλότερο ρυθμό στο αναπτυσσόμενο (+) άκρο. Αλλά εάν η συγκέντρωση της τουμπουλίνης είναι ανεπαρκής, οι μικροσωληνίσκοι μπορούν να αποσυναρμολογηθούν και στα δύο άκρα. Η αποσυναρμολόγηση των μικροσωληνίσκων διευκολύνεται από τη μείωση της θερμοκρασίας και την παρουσία ιόντων Ca ++.
Οι μικροσωληνίσκοι είναι πολύ δυναμικές δομές που μπορούν να προκύψουν και να αποσυναρμολογηθούν αρκετά γρήγορα. Οι απομονωμένοι μικροσωληνίσκοι περιέχουν πρόσθετες πρωτεΐνες που σχετίζονται με αυτά, τα λεγόμενα. MAP πρωτεΐνες (MAP - microtubule accessory proteins). Αυτές οι πρωτεΐνες, σταθεροποιώντας τους μικροσωληνίσκους, επιταχύνουν τη διαδικασία του πολυμερισμού της τουμπουλίνης (Εικ. 269).
Ο ρόλος των κυτταροπλασματικών μικροσωληνίσκων περιορίζεται στην εκτέλεση δύο λειτουργιών: του σκελετικού και του κινητικού. Ο σκελετικός ρόλος του πλαισίου είναι ουσιαστικά ότι η διάταξη των μικροσωληνίσκων στο κυτταρόπλασμα σταθεροποιεί το σχήμα του κυττάρου. Όταν οι μικροσωληνίσκοι διαλύονται, τα κύτταρα που είχαν πολύπλοκο σχήμα τείνουν να αποκτούν σφαιρικό σχήμα. Ο κινητικός ρόλος των μικροσωληνίσκων δεν έγκειται μόνο στο γεγονός ότι δημιουργούν ένα διατεταγμένο, διανυσματικό σύστημα κίνησης. Οι κυτταροπλασματικοί μικροσωληνίσκοι, σε συνδυασμό με συγκεκριμένες σχετιζόμενες κινητικές πρωτεΐνες, σχηματίζουν σύμπλοκα ATPase που μπορούν να οδηγήσουν κυτταρικά συστατικά.
Σχεδόν σε όλα τα ευκαρυωτικά κύτταρα, στο υαλόπλασμα μπορούν να παρατηρηθούν μακροί, μη διακλαδιζόμενοι μικροσωληνίσκοι. Βρίσκονται σε μεγάλες ποσότητες στις κυτταροπλασματικές διεργασίες των νευρικών κυττάρων, στις διεργασίες των μελανοκυττάρων, των αμοιβάδων και άλλων κυττάρων που αλλάζουν το σχήμα τους (Εικ. 270). Μπορούν να απομονωθούν οι ίδιοι ή να απομονωθούν οι πρωτεΐνες τους: πρόκειται για τις ίδιες τουμπουλίνες με όλες τις ιδιότητές τους.
Κέντρα οργάνωσης μικροσωληνίσκων.Η ανάπτυξη των μικροσωληνίσκων στο κυτταρόπλασμα συμβαίνει πολικά: το (+) άκρο του μικροσωληνίσκου μεγαλώνει. Η διάρκεια ζωής των μικροσωληνίσκων είναι πολύ μικρή, και ως εκ τούτου δημιουργούνται συνεχώς νέοι μικροσωληνίσκοι. Η διαδικασία της έναρξης του πολυμερισμού της τουμπουλίνης, η πυρήνωση, συμβαίνει σε σαφώς καθορισμένες περιοχές του κυττάρου, στο λεγόμενο. κέντρα οργάνωσης μικροσωληνίσκων (MTOCs). Στις ζώνες COMMT γίνεται η τοποθέτηση κοντών μικροσωληνίσκων, με τα (-) άκρα τους στραμμένα προς το COMMT. Πιστεύεται ότι στις ζώνες COMT (--) τα άκρα μπλοκάρονται από ειδικές πρωτεΐνες που εμποδίζουν ή περιορίζουν τον αποπολυμερισμό των τουμπουλινών. Για το λόγο αυτό, με επαρκή ποσότητα ελεύθερης τουμπουλίνης, το μήκος των μικροσωληνίσκων που εκτείνονται από το COMT θα αυξηθεί. Κυρίως κυτταρικά κέντρα που περιέχουν κεντριόλια συμμετέχουν ως COMMT σε ζωικά κύτταρα, όπως θα συζητηθεί παρακάτω. Επιπλέον, η πυρηνική ζώνη, και κατά τη διάρκεια της μίτωσης, οι πόλοι της ατράκτου, μπορούν να χρησιμεύσουν ως COMMT.
Ένας από τους σκοπούς των κυτταροπλασματικών μικροσωληνίσκων είναι να δημιουργήσουν έναν ελαστικό, αλλά ταυτόχρονα σταθερό ενδοκυτταρικό σκελετό, ο οποίος είναι εξαιρετικά σημαντικός για τη διατήρηση του σχήματος του κυττάρου. Στα αμφίβια ερυθροκύτταρα σε σχήμα δίσκου, μια δέσμη κυκλικά διατεταγμένων μικροσωληνίσκων βρίσκεται κατά μήκος της περιφέρειας του κυττάρου. δέσμες μικροσωληνίσκων είναι χαρακτηριστικές για διάφορες εκβολές του κυτταροπλάσματος (αξοπόδια πρωτοζώων, άξονες νευρικών κυττάρων κ.λπ.).
Ο ρόλος των μικροσωληνίσκων είναι να σχηματίσουν ένα πλαίσιο για την υποστήριξη του κυτταρικού σώματος, τη σταθεροποίηση και την ενίσχυση των κυτταρικών αποβλήτων. Ταυτόχρονα, οι μικροσωληνίσκοι εμπλέκονται στις διαδικασίες ανάπτυξης των κυττάρων. Έτσι, στα φυτά, κατά τη διαδικασία της επιμήκυνσης των κυττάρων, όταν λόγω αύξησης του κεντρικού κενοτόπιου, συμβαίνει σημαντική αύξηση του όγκου των κυττάρων, εμφανίζονται μεγάλοι αριθμοί μικροσωληνίσκων στα περιφερειακά στρώματα του κυτταροπλάσματος. Σε αυτή την περίπτωση, οι μικροσωληνίσκοι, καθώς και το κυτταρικό τοίχωμα που αναπτύσσεται αυτή τη στιγμή, φαίνεται να ενισχύουν και να ενισχύουν μηχανικά το κυτταρόπλασμα.
Με τη δημιουργία του ενδοκυτταρικού σκελετού, οι μικροσωληνίσκοι αποτελούν παράγοντες στην προσανατολισμένη κίνηση των ενδοκυτταρικών συστατικών, θέτοντας με τη διάταξή τους χώρους για κατευθυνόμενες ροές διαφόρων ουσιών και για την κίνηση μεγάλων δομών.
Δημοσιεύτηκε στο ref.rf
Έτσι, στην περίπτωση των μελανοφόρων (κύτταρα που περιέχουν τη χρωστική μελανίνη) των ψαριών, όταν αναπτύσσονται οι κυτταρικές διεργασίες, οι κόκκοι χρωστικής κινούνται κατά μήκος δεσμών μικροσωληνίσκων.
Στους άξονες των ζωντανών νευρικών κυττάρων, μπορεί κανείς να παρατηρήσει την κίνηση διαφόρων μικρών κενοτοπίων και κόκκων, τα οποία κινούνται τόσο από το σώμα του κυττάρου προς τη νευρική απόληξη (προσθοριακή μεταφορά) όσο και προς την αντίθετη κατεύθυνση (ανάδρομη μεταφορά).
Απομονώθηκαν πρωτεΐνες που είναι υπεύθυνες για την κίνηση των κενοτοπίων. Ένα από αυτά είναι η κινεσίνη, μια πρωτεΐνη με μοριακό βάρος περίπου 300 χιλιάδες.
Υπάρχει μια ολόκληρη οικογένεια κινεσινών. Έτσι, οι κυτοσολικές κινεσίνες εμπλέκονται στη μεταφορά κυστιδίων, λυσοσωμάτων και άλλων μεμβρανικών οργανιδίων κατά μήκος των μικροσωληνίσκων. Πολλές από τις κινεσίνες συνδέονται ειδικά με τα φορτία τους. Έτσι, μερικοί εμπλέκονται στη μεταφορά μόνο μιτοχονδρίων, άλλοι - μόνο συναπτικών κυστιδίων. Οι κινεσίνες συνδέονται με τις μεμβράνες μέσω συμπλεγμάτων πρωτεϊνών μεμβράνης - κινεκτινών. Οι κινεσίνες της ατράκτου εμπλέκονται στο σχηματισμό αυτής της δομής και στην απόκλιση των χρωμοσωμάτων.
Μια άλλη πρωτεΐνη, η κυτταροπλασματική δυνεΐνη, είναι υπεύθυνη για την ανάδρομη μεταφορά στον άξονα (Εικ. 275). Αποτελείται από δύο βαριές αλυσίδες - κεφαλές που αλληλεπιδρούν με μικροσωληνίσκους, αρκετές ενδιάμεσες και ελαφριές αλυσίδες που συνδέονται με μεμβρανικά κενοτόπια. Η κυτταροπλασματική δυνεΐνη είναι μια κινητήρια πρωτεΐνη που μεταφέρει φορτίο στο μείον άκρο των μικροσωληνίσκων. Οι δυνεΐνες χωρίζονται επίσης σε δύο κατηγορίες: κυτοσολικές - που εμπλέκονται στη μεταφορά κενοτοπίων και χρωμοσωμάτων, και αξονικές - υπεύθυνες για την κίνηση των βλεφαρίδων και των μαστιγίων.
Κυτοπλασματικές δυνεΐνες και κινεσίνες έχουν βρεθεί σχεδόν σε όλους τους τύπους ζωικών και φυτικών κυττάρων.
Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, και στο κυτταρόπλασμα, η κίνηση πραγματοποιείται σύμφωνα με την αρχή των συρόμενων νημάτων, μόνο που δεν κινούνται νήματα κατά μήκος μικροσωληνίσκων, αλλά μικρά μόρια - κινητές που σχετίζονται με κινούμενα κυτταρικά συστατικά. Η ομοιότητα με το σύμπλεγμα ακτομυοσίνης αυτού του συστήματος ενδοκυτταρικής μεταφοράς είναι ουσιαστικά ότι σχηματίζεται ένα διπλό σύμπλεγμα (μικροσωληνίσκος + κινητήριος), το οποίο έχει υψηλή δραστηριότητα ΑΤΡάσης.
Όπως φαίνεται, οι μικροσωληνίσκοι σχηματίζουν ακτινικά αποκλίνοντα πολωμένα ινίδια στο κύτταρο, τα (+) άκρα των οποίων κατευθύνονται από το κέντρο του κυττάρου προς την περιφέρεια. Η παρουσία (+) και (-)-κατευθυνόμενων πρωτεϊνών κινητήρα (κινεσίνες και δινεΐνες) δημιουργεί την ευκαιρία για τη μεταφορά των συστατικών τους στο κύτταρο τόσο από την περιφέρεια στο κέντρο (ενδοκυττατικά κενοτόπια, ανακύκλωση κενοτοπίων ER και συσκευή Golgi κ.λπ.) και από το κέντρο προς την περιφέρεια (ER vacuoles, λυσοσώματα, εκκριτικά vacuoles κ.λπ.) (Εικ. 276). Αυτή η πολικότητα μεταφοράς δημιουργείται λόγω της οργάνωσης ενός συστήματος μικροσωληνίσκων που προκύπτουν στα κέντρα της οργάνωσής τους, στο κυτταρικό κέντρο.
Μικροσωληνίσκοι - έννοια και τύποι. Ταξινόμηση και χαρακτηριστικά της κατηγορίας "Μικροσωληνίσκοι" 2017, 2018.
Στα κύτταρα, οι μικροσωληνίσκοι συμμετέχουν στη δημιουργία ενός αριθμού προσωρινών (κυτταροσκελετός κυττάρων μεσοφάσης, ατράκτης) ή μόνιμων (κεντρόλια, βλεφαρίδες, μαστίγια) δομές.
Οι μικροσωληνίσκοι είναι ίσιοι, μη διακλαδιζόμενοι, μακροί, κοίλοι κύλινδροι (βλ. Εικ. 18). Η εξωτερική τους διάμετρος είναι περίπου 24 nm, ο εσωτερικός αυλός έχει πλάτος 15 nm και το πάχος τοιχώματος είναι 5 nm. Το τοίχωμα του μικροσωληνίσκου είναι κατασκευασμένο από πυκνά γεμάτες στρογγυλές υπομονάδες με διάμετρο περίπου 5 nm. Σε ένα ηλεκτρονικό μικροσκόπιο, οι διατομές των μικροσωληνίσκων δείχνουν ως επί το πλείστον 13 υπομονάδες διατεταγμένες σε ένα δακτύλιο μονής στρώσης. Οι μικροσωληνίσκοι που απομονώνονται από διαφορετικές πηγές (πρωτόζωα βλεφαρίδες, κύτταρα νευρικού ιστού, άτρακτοι) έχουν παρόμοια σύνθεση και περιέχουν πρωτεΐνες - τουμπουλίνες. Σχεδόν σε όλα τα ευκαρυωτικά κύτταρα, στο υαλόπλασμα μπορούν να παρατηρηθούν μακροί, μη διακλαδιζόμενοι μικροσωληνίσκοι. Βρίσκονται σε μεγάλες ποσότητες στις κυτταροπλασματικές διεργασίες των νευρικών κυττάρων, των ινοβλαστών και άλλων κυττάρων που αλλάζουν το σχήμα τους.
Μία από τη λειτουργική σημασία τέτοιων κυτταροπλασματικών μικροσωληνίσκων είναι να δημιουργήσουν ένα ελαστικό, αλλά ταυτόχρονα σταθερό ενδοκυτταρικό πλαίσιο (κυτταροσκελετός), απαραίτητο για τη διατήρηση του σχήματος του κυττάρου.
Δημιουργώντας έναν ενδοκυτταρικό σκελετό, οι μικροσωληνίσκοι μπορούν να είναι παράγοντες στην προσανατολισμένη κίνηση του κυττάρου στο σύνολό του και των ενδοκυττάριων συστατικών του, και με τη διάταξη τους, ορίζουν φορείς για κατευθυνόμενες ροές διαφόρων ουσιών και για την κίνηση μεγάλων δομών.
Η καταστροφή των μικροσωληνίσκων από την κολχικίνη διαταράσσει τη μεταφορά ουσιών στους άξονες των νευρικών κυττάρων, οδηγεί σε αποκλεισμό της έκκρισης κ.λπ.
9. Λυσοσώματα: δομή, λειτουργίες, ταξινόμηση
Τα λυσοσώματα είναι μια διαφορετική κατηγορία κενοτοπίων 0,2-0,4 μm που οριοθετούνται από μία μόνο μεμβράνη. Χαρακτηριστικό γνώρισμα των λυσοσωμάτων είναι η παρουσία σε αυτά υδρολυτικών ενζύμων - υδρολασών (πρωτεϊνάσες, νουκλεάσες, γλυκοσιδάσες, φωσφατάσες, λιπάσες), που διασπούν διάφορα βιοπολυμερή σε όξινο pH. Τα λυσοσώματα ανακαλύφθηκαν το 1949 από τον de Duve.
Μεταξύ των λυσοσωμάτων, διακρίνονται τουλάχιστον 3 τύποι: πρωτογενή λυσοσώματα, δευτερογενή λυσοσώματα (φαγολυσοσώματα και αυτοφαγοσώματα) και υπολειμματικά σώματα. Η ποικιλομορφία της μορφολογίας του λυσοσώματος εξηγείται από το γεγονός ότι αυτά τα σωματίδια συμμετέχουν στις διαδικασίες της ενδοκυτταρικής πέψης, σχηματίζοντας πολύπλοκα πεπτικά κενοτόπια τόσο εξωγενούς (εξωκυτταρικής) όσο και ενδογενούς (ενδοκυτταρικής) προέλευσης.
Τα πρωτογενή λυσοσώματα είναι μικρά κυστίδια μεμβράνης μεγέθους περίπου 0,2-0,5 μm, γεμάτα με μια ουσία χωρίς δομή που περιέχει υδρολάσες, συμπεριλαμβανομένης της δραστικής όξινης φωσφατάσης, η οποία είναι ένα ένζυμο δείκτη για τα λυσοσώματα. Αυτά τα μικρά κυστίδια είναι σχεδόν πολύ δύσκολο να διακριθούν από τα μικρά κυστίδια στην περιφέρεια της συσκευής Golgi, τα οποία περιέχουν επίσης όξινη φωσφατάση. Η θέση της σύνθεσής του είναι το κοκκώδες ενδοπλασματικό δίκτυο.
Τα δευτερογενή λυσοσώματα, ή τα ενδοκυτταρικά πεπτικά κενοτόπια, σχηματίζονται από τη σύντηξη πρωτογενών λυσοσωμάτων με φαγοκυτταρικά ή πινοκυτταρωτικά κενοτόπια, σχηματίζοντας φαγολυσοσώματα ή ετεροφαγοσώματα, καθώς και με τροποποιημένα οργανίδια του ίδιου του κυττάρου που υφίστανται πέψη. Οι ουσίες που εισέρχονται στο δευτερογενές λυσόσωμα διασπώνται από υδρολάσες σε μονομερή, τα οποία μεταφέρονται μέσω της μεμβράνης του λυσοσώματος στο υαλόπλασμα, όπου επαναχρησιμοποιούνται, δηλ. περιλαμβάνονται σε διάφορες μεταβολικές διεργασίες.
Ωστόσο, η διάσπαση και η πέψη των βιογενών μακρομορίων μέσα στα λυσοσώματα μπορεί να μην ολοκληρωθεί σε ορισμένα κύτταρα. Σε αυτή την περίπτωση, τα άπεπτα προϊόντα συσσωρεύονται στις κοιλότητες των λυσοσωμάτων. Αυτό το λυσόσωμα ονομάζεται τελολυσόσωμα ή υπολειπόμενο σώμα. Τα υπολειμματικά σώματα περιέχουν λιγότερα υδρολυτικά ένζυμα, τα περιεχόμενα συμπιέζονται και αναδιατάσσονται. Για παράδειγμα, στους ανθρώπους, καθώς το σώμα γερνά, η «χρωστική ουσία γήρανσης» - λιποφουσκίνη - εναποτίθεται στα κύτταρα του εγκεφάλου, του ήπατος και στις μυϊκές ίνες στα τελολυσοσώματα.
Η λειτουργική σημασία της αυτοφαγοκυττάρωσης είναι ακόμα ασαφής. Υπάρχει η υπόθεση ότι αυτή η διαδικασία σχετίζεται με την επιλογή και την καταστροφή αλλοιωμένων, κατεστραμμένων κυτταρικών συστατικών. Σε αυτή την περίπτωση, τα λυσοσώματα λειτουργούν ως ενδοκυτταρικά «καθαριστικά» που αφαιρούν τις ελαττωματικές δομές.
Σχεδόν σε όλα τα ευκαρυωτικά κύτταρα στο υαλόπλασμα μπορεί κανείς να δει μακριά, μη διακλαδισμένα μικροσωληνίσκους. Βρίσκονται σε μεγάλες ποσότητες στις κυτταροπλασματικές διεργασίες των νευρικών κυττάρων, των ινοβλαστών και άλλων κυττάρων που αλλάζουν το σχήμα τους. Μπορούν να απομονωθούν οι ίδιοι ή να απομονωθούν οι πρωτεΐνες που τις σχηματίζουν: πρόκειται για τις ίδιες τουμπουλίνες με όλες τις ιδιότητές τους.
Κύρια λειτουργική αξίαΟ σκοπός τέτοιων κυτταροπλασματικών μικροσωληνίσκων είναι να δημιουργήσουν ένα ελαστικό, αλλά ταυτόχρονα σταθερό ενδοκυτταρικό πλαίσιο (κυτταροσκελετός), απαραίτητο για τη διατήρηση του σχήματος του κυττάρου.
Τα οργανίδια μιας δομής χωρίς μεμβράνη περιλαμβάνουν μικροσωληνίσκους - σωληνοειδείς δομές διαφόρων μηκών με εξωτερική διάμετρο 24 nm, πάχος τοιχώματος περίπου 5 nm και πλάτος "αυλού" 15 nm. Βρίσκονται ελεύθερα στο κυτταρόπλασμα των κυττάρων ή ως δομικά στοιχεία των μαστιγίων (σπερματοζωάρια), των βλεφαρίδων (κυλιωμένο επιθήλιο της τραχείας), της μιτωτικής ατράκτου και των κεντρολίων (διαιρούμενα κύτταρα).
Οι μικροσωληνίσκοι κατασκευάζονται από τη συναρμολόγηση (πολυμερισμό) της πρωτεϊνικής τουμπουλίνης. Μικροσωληνίσκοι πολικός:έχουν τα άκρα (+) και (-). Η ανάπτυξή τους προέρχεται από την ειδική δομή των μη διαιρούμενων κυττάρων - κέντρο οργάνωσης μικροσωληνίσκων, με το οποίο συνδέεται το οργανίδιο με το άκρο (-) και το οποίο αντιπροσωπεύεται από δύο στοιχεία πανομοιότυπα σε δομή με τα κεντρόλια του κυτταρικού κέντρου. Η επιμήκυνση των μικροσωληνίσκων συμβαίνει από προσάρτηση νέων υπομονάδων στο τέλος (+).Στην αρχική φάση δεν προσδιορίζεται η κατεύθυνση ανάπτυξης, αλλά από τους μικροσωληνίσκους που προκύπτουν διατηρούνται αυτοί που έρχονται σε επαφή με το (+) άκρο τους με κατάλληλο στόχο. Σε φυτικά κύτταρα που έχουν μικροσωληνίσκους, δεν έχουν βρεθεί δομές τύπου κεντρολίου.
Οι μικροσωληνίσκοι συμμετέχουν:
- στη διατήρηση του σχήματος των κυττάρων,
- στην οργάνωση της κινητικής τους δραστηριότητας (μαστίγια, βλεφαρίδες) και της ενδοκυτταρικής μεταφοράς (χρωμοσώματα σε ανάφαση της μίτωσης).
Οι λειτουργίες των ενδοκυτταρικών μοριακών κινητήρων εκτελούνται από τις πρωτεΐνες κινεσίνη και dynein, οι οποίες έχουν τη δραστηριότητα του ενζύμου ATPase. Κατά τη διάρκεια της μαστιγωτής ή βλεφαριδωτής κίνησης, τα μόρια της dynein, προσκολλώνται σε μικροσωληνίσκους και χρησιμοποιώντας την ενέργεια του ATP, κινούνται κατά μήκος της επιφάνειάς τους προς το βασικό σώμα, δηλαδή προς το άκρο (-). Η μετατόπιση των μικροσωληνίσκων μεταξύ τους προκαλεί κυματοειδείς κινήσεις του μαστιγίου ή των βλεφαρίδων, ωθώντας το κύτταρο να κινηθεί στο διάστημα. Στην περίπτωση των ακίνητων κυττάρων, για παράδειγμα, του βλεφαροφόρου επιθηλίου της τραχείας, ο περιγραφόμενος μηχανισμός χρησιμοποιείται για την απομάκρυνση της βλέννας από την αναπνευστική οδό με τα σωματίδια να καθιζάνουν σε αυτήν (λειτουργία παροχέτευσης).
Η συμμετοχή των μικροσωληνίσκων στην οργάνωση της ενδοκυτταρικής μεταφοράς απεικονίζει την κίνηση των κυστιδίων (κυστιδίων) στο κυτταρόπλασμα. Τα μόρια κινεσίνης και δυνεΐνης περιέχουν δύο σφαιρικές «κεφαλές» και «ουρές» με τη μορφή πρωτεϊνικών αλυσίδων. Με τη βοήθεια των κεφαλών τους, οι πρωτεΐνες έρχονται σε επαφή με μικροσωληνίσκους, κινούμενοι κατά μήκος της επιφάνειάς τους: κινεσίνη από άκρο (-) σε άκρο (+) και δυνεΐνη προς την αντίθετη κατεύθυνση. Ταυτόχρονα, τραβούν πίσω τους φυσαλίδες που συνδέονται με τις «ουρές» τους. Προφανώς, η μακρομοριακή οργάνωση των «ουρών» είναι μεταβλητή, γεγονός που εξασφαλίζει την αναγνώριση των διαφόρων μεταφερόμενων δομών.
Οι μικροσωληνίσκοι, ως βασικό συστατικό της μιτωτικής συσκευής, σχετίζονται με την απόκλιση των κεντρολίων προς τους πόλους ενός διαιρούμενου κυττάρου και την κίνηση των χρωμοσωμάτων στην ανάφαση της μίτωσης. Τα ζωικά κύτταρα, τα κύτταρα των μερών των φυτών, οι μύκητες και τα φύκια χαρακτηρίζονται από ένα κυτταρικό κέντρο (διπλόσωμα), που σχηματίζεται από δύο κεντρόλες. Κάτω από ένα ηλεκτρονικό μικροσκόπιο, το κεντριόλιο μοιάζει με έναν «κούφιο» κύλινδρο με διάμετρο 150 nm και μήκος 300-500 nm. Το τοίχωμα του κυλίνδρου σχηματίζεται από 27 μικροσωληνίσκους, ομαδοποιημένους σε 9 τρίδυμα. Η λειτουργία των κεντρολίων, παρόμοιας δομής με τα στοιχεία του κέντρου οργάνωσης μικροσωληνίσκων (δείτε εδώ, παραπάνω), περιλαμβάνει το σχηματισμό νημάτων μιτωτικής ατράκτου (άτρακτος διαίρεσης, άτρακτος αχρωματίνης κλασικής κυτταρολογίας), που είναι μικροσωληνίσκοι. Τα κεντρόλια πολώνουν τη διαδικασία της κυτταρικής διαίρεσης, διασφαλίζοντας τη φυσική απόκλιση των αδελφών χρωματίδων (θυγατρικών χρωμοσωμάτων) στους πόλους τους στην ανάφαση της μίτωσης
Δομή κινεσίνης (α) και μεταφορά κυστιδίων κατά μήκος μικροσωληνίσκου (β)
Γύρω από κάθε κεντριόλιο υπάρχει μια μήτρα χωρίς δομή ή λεπτό ινώδη. Μπορείτε συχνά να βρείτε πολλές πρόσθετες δομές που σχετίζονται με κεντρόλια: δορυφόρους, εστίες σύγκλισης μικροσωληνίσκων, πρόσθετους μικροσωληνίσκους που σχηματίζουν μια ειδική ζώνη, μια κεντρόσφαιρα γύρω από το κεντριόλιο.
Οι μικροσωληνίσκοι βρίσκονται, κατά κανόνα, στα βαθύτερα στρώματα του κυτοσόλης που βρίσκεται κοντά στη μεμβράνη. Επομένως, οι περιφερειακοί μικροσωληνίσκοι πρέπει να θεωρούνται ως μέρος του δυναμικού, οργανωτικού μικροσωληνίσκου «σκελετού» του κυττάρου. Ωστόσο, τόσο οι συσταλτικές όσο και οι σκελετικές ινιδώδεις δομές του περιφερικού κυτταρολύματος συνδέονται επίσης άμεσα με τις ινώδεις δομές του κύριου υαλοπλάσματος του κυττάρου. Λειτουργικά, το περιφερειακό στήριγμα-συσταλτικό ινιδιακό σύστημα του κυττάρου βρίσκεται σε στενή αλληλεπίδραση με το σύστημα των περιφερειακών μικροσωληνίσκων. Αυτό μας δίνει λόγο να θεωρήσουμε το τελευταίο ως μέρος του υπομεμβρανικού συστήματος του κυττάρου.
Το σύστημα μικροσωληνίσκων είναι το δεύτερο συστατικό της συσκευής στήριξης-συστολής, η οποία είναι, κατά κανόνα, σε στενή επαφή με το μικροϊνίδιο εξάρτημα. Τα τοιχώματα των μικροσωληνίσκων σχηματίζονται στη διατομή τις περισσότερες φορές από 13 διμερή πρωτεϊνικά σφαιρίδια, κάθε σφαιρίδιο αποτελείται από α- και β-τουμπουλίνες (Εικ. 6). Οι τελευταίοι στους περισσότερους μικροσωληνίσκους είναι διατεταγμένοι σε μοτίβο σκακιέρας. Η τουμπουλίνη αποτελεί το 80% των πρωτεϊνών που περιέχονται στους μικροσωληνίσκους. Το υπόλοιπο 20% αντιστοιχεί στις πρωτεΐνες υψηλού μοριακού βάρους MAP 1, MAP 2 και χαμηλού μοριακού βάρους παράγοντα tau. Οι πρωτεΐνες MAP (πρωτεΐνες που σχετίζονται με μικροσωληνίσκους) και ο παράγοντας tau είναι συστατικά απαραίτητα για τον πολυμερισμό της τουμπουλίνης. Ελλείψει αυτών, η αυτοσυναρμολόγηση των μικροσωληνίσκων μέσω του πολυμερισμού της τουμπουλίνης είναι εξαιρετικά δύσκολη και οι μικροσωληνίσκοι που προκύπτουν είναι πολύ διαφορετικοί από τους εγγενείς.
Οι μικροσωληνίσκοι είναι μια πολύ ασταθής δομή, για παράδειγμα, οι μικροσωληνίσκοι θερμόαιμων ζώων καταστρέφονται συνήθως στο κρύο. Υπάρχουν επίσης μικροσωληνίσκοι ανθεκτικοί στο κρύο, για παράδειγμα, στους νευρώνες του κεντρικού νευρικού συστήματος των σπονδυλωτών, ο αριθμός τους κυμαίνεται από 40 έως 60%. Οι θερμοσταθεροί και οι θερμοευκίνητοι μικροσωληνίσκοι δεν διαφέρουν ως προς τις ιδιότητες της τουμπουλίνης που περιέχουν. Προφανώς, αυτές οι διαφορές καθορίζονται από πρόσθετες πρωτεΐνες. Στα εγγενή κύτταρα, σε σύγκριση με τα μικροϊνίδια, το κύριο μέρος του υπομεμβρανικού συστήματος των μικροσωληνίσκων βρίσκεται σε βαθύτερες περιοχές του κυτταροπλάσματος Υλικό από τον ιστότοπο
Ακριβώς όπως τα μικροϊνίδια, οι μικροσωληνίσκοι υπόκεινται σε λειτουργική μεταβλητότητα. Χαρακτηρίζονται από αυτοσυναρμολόγηση και αυτοαποσυναρμολόγηση, με την αποσυναρμολόγηση να συμβαίνει μέχρι τα διμερή τουμπουλίνης. Συνεπώς, οι μικροσωληνίσκοι μπορούν να αναπαρασταθούν σε μεγαλύτερο ή μικρότερο αριθμό λόγω της κυριαρχίας των διαδικασιών είτε αυτο-αποσυναρμολόγησης είτε αυτοσυναρμολόγησης μικροσωληνίσκων από το ταμείο της σφαιρικής τουμπουλίνης του υαλοπλάσματος. Οι έντονες διαδικασίες αυτοσυναρμολόγησης μικροσωληνίσκων συνήθως περιορίζονται σε θέσεις κυτταρικής προσκόλλησης στο υπόστρωμα, δηλ. σε θέσεις ενισχυμένου πολυμερισμού της ινιδιακής ακτίνης από τη σφαιρική ακτίνη του υαλοπλάσματος. Αυτή η συσχέτιση του βαθμού ανάπτυξης αυτών των δύο μηχανοχημικών συστημάτων δεν είναι τυχαία και αντανακλά τη βαθιά λειτουργική τους σχέση σε ολόκληρο το μυοσκελετικό και μεταφορικό σύστημα του κυττάρου.
- Σε επαφή με 0
- Google+ 0
- Εντάξει 0
- Facebook 0